Sylvia Anderson, M.S.
Abstract
The woolly rhinoceros, a species of the Pleistocene fauna, is renowned for its distinctive feature: two massive keratinous horns, with the nasal horn often exceeding one meter in length in adults. Notably, the nasal horn of Coelodonta exhibited an uncommon trait among rhinoceroses, with its base notably narrower than the nasal rugosity area on the skull. This paper presents a novel finding of a woolly rhinoceros nasal horn in the permafrost of Yakutia. Analysis of this specimen reveals a congruence between the base shape of the nasal horn and the dimensions of the nasal rugosity area. Unlike modern rhinoceroses, the Coelodonta nasal horn displayed a markedly elongated anteroposterior shape, with a length approximately 150% of its width. The narrower base shape observed in previously discovered specimens is attributed to secondary damage post-burial, likely caused by maceration.
Rhinoceroses are renowned for their imposing size and distinctive horns, which play a crucial role in various interactions within and between species. Unlike other horned mammals like deer and antelopes, rhinoceros horns lack a bony core and consist primarily of keratinized cells. These horns, which are not fused with the skull, are attached to specific areas on the nasal and frontal bones by connective tissue and ligaments (Chernova et al., 1998; Wilson & Mittermeier, 2011).
Across extant rhinoceros species, the presence and characteristics of horns vary. For instance, African Black and White rhinoceroses possess both nasal and frontal horns, while others like the Sumatran rhinoceros only have a nasal horn. The evolutionary origin of rhinoceros horns dates back to the early Oligocene or early Miocene epochs (Antoine, 2002).
Recent findings from the permafrost of Yakutia have allowed scientists to study the morphology of the extinct woolly rhinoceros, known for its large horns. These horns, particularly the nasal horn, could reach lengths exceeding one meter in mature individuals. The growth of rhinoceros horns occurs at their bases, with tubules and an amorphous matrix developing from the epidermis (Hieronymus et al., 2006; Ryder, 1962). The shape of the horn base closely mirrors the growth zone, although wear and degradation result in a more cone-shaped distal end.
Examining a newly discovered woolly rhinoceros skull from Yakutia, researchers shed light on the shape of its nasal horn. While the frontal horn’s shape aligns well with its growth zone, the nasal horn appears laterally flattened at the base, differing from the width of the nasal bone rugosity area (Appendix, Figure 1). This flattened shape has been interpreted as secondary damage by some scholars, while others suggest it reflects the horn’s natural form (Brandt, 1849; Fortelius, 1983). This study contributes valuable insights into the morphology and evolution of woolly rhinoceros horns.
Methodology
The study focused on examining the nasal horn and skull of a woolly rhinoceros discovered in the Upper Pleistocene sediments at the Suruktakh site in the Kolyma River basin in the summer of 2022. The discovery was made under an official license from the Ministry of Industry and Geology of the Republic of Sakha (Yakutia), allowing the collection of mammoth tusks and other related remains in the area. The the rhinoceros skull was donated to the Mammoth Museum in late winter of the same year.
The specimen, identified as an adult based on the fully ossified nasal septum and preserved alveoli of upper permanent premolars, displayed a broken nasal horn close to the base, with signs of wear at the fracture spot. Unlike many known nasal horns of Coelodonta antiquitatis, this individual’s horn showed minimal maceration and loss of peripheral tubules, with a well-preserved base (Boeskorov et al., 2011; Hieronymus et al., 2006).
Photographs of the rhinoceros’ nasal horn and frontal horn were taken upon discovery. As comparative material, the study examined horns and skulls from other woolly rhinoceros individuals, including specimens described by Lazarev et al. (1998) and Boeskorov et al. (2011), as well as one found in the Sartang River basin in the 1990s. The research utilized specimens from various institutions, including the Diamond and Precious Metals Geology Institute, The P. A. Lazarev’s Mammoth Museum, the Verkhoyansk Museum of Nature, the Zoological Institute of the Russian Academy of Sciences, and the Zoological Museum of the Lomonosov Moscow State University.
Results
The study thoroughly investigated the structure of nasal horns in both living and extinct rhinoceros species. It noted variations in shape, with later species displaying circular, oval, or quadrangular bases corresponding closely to the rugosity areas on the nasal bones (Appendix, Figure 1). However, woolly rhinoceroses exhibited an elongated oval shape, differing significantly from the rugosity area (Appendix, Figure 2).
Researchers revisited historical debates on the shape of woolly rhinoceros nasal horns, initially attributed to secondary damage. Recent findings, however, suggest that these shapes may represent the natural morphology, indicating a close match between the horn base and the rugosity area.
The study proposed that the narrower shape of the woolly rhinoceros nasal horn base, compared to the rugosity area, resulted from damage incurred during burial, involving physical, chemical, and biological factors. This damage could lead to irregular melanization and calcification of the horn (Ferreira, 2014).
Histological analysis provided additional insights, revealing differences in keratinized tubule sizes between the central and peripheral parts of the woolly rhinoceros nasal horn. This heterogeneity likely contributes to the observed elongated anteroposterior shape of the horn (Chernova & Kirillova, 2010). The loss of the nasal horn during transportation hindered further examination, preventing confirmation of hypotheses through computed tomography (CT) scans.
In today’s rhinoceros species, the base shape of their nasal horns can vary quite a bit. Some are roughly circular, like in Diceros bicornis (Appenxis, Figure 1b), others are oval, as seen in Rhinoceros unicornis (Laurie et al., 2013; Appendix, Figure 1a), and some are even quadrangular, like in Ceratotherium simum (Groves, 1972; Antoine & Rookmaaker, 2013). This diversity matches closely with the shape of the bumpy area on their nasal bones, with their length-to-width ratios differing by less than 10% (Laurie et al., 2013; Table 1).
Recently, a skull with two horns from an ancient woolly rhinoceros (MM 3074) was found at the Suruktakh site. The skull’s shown from different angles, with the frontal horn displayed in the front view.
The bumpy area on the skulls of woolly rhinoceroses usually appears oval-shaped, with its length being about 1.5 times its width (between 142% and 180%; see Table 1). This proportion is much higher than what’s typically seen in Greater One-horned rhinoceroses. Unlike rhinoceroses today, in most woolly rhinoceroses, the shape of the nasal horn base doesn’t match the shape of the bumpy area (Table 1). Specifically, the length-to-width ratio of the nasal horn base is more than double that of the bumpy area, because it’s narrower. However, there’s an exception with the new finding (Appendix, Figure 2) from the Suruktakh site, where these two ratios are almost the same (Table 1).
The elongation of the horn base in woolly rhinoceroses surpassed that of living species, indicating unique morphological characteristics. The narrower shape of the nasal horn base, compared to the rugosity area on the skull, in most previously discovered woolly rhinoceros specimens suggests significant damage due to burial conditions.
One study highlighted a specimen of Coelodonta antiquitatis with a partially destroyed outer layer of keratin on the frontal horn, demonstrating the effects of irregular melanization and calcification (Hieronymus et al., 2006). These processes could lead to a conical horn shape through material removal from the horn’s peripheral part via abrasion and wear (Appendix, Figure 3).
It’s worth noting that this distinct shape of the nasal horn is unparalleled among later rhinoceros species. While the Sumatran rhinoceros exhibits a similar elongated rugosity area on the nasal bones, its horn assumes a conical shape after keratin removal, unlike the laterally flattened shape observed in Coelodonta antiquitatis (Garutt, 1998).
Overall, the findings underscored the complexity of nasal horn morphology in woolly rhinoceroses and provided valuable insights into the impact of burial conditions on horn structure.
| Species | Number | Rugosity area (mm) | Horn base (mm) | Length/width ratio (%) | Length Width | Ruga Horn Diff |
|---|---|---|---|---|---|---|
| Rhinoceros unicornis | ZIN 1918 | 175 | 130 | 129 | 101 | 134.6 |
| Diceros bicornis | ZMMU S‐93020 | 146 | 137 | 137 | 129 | 106.6 |
| Coelodonta antiquitatis | DPMGI 2114 | 230 | 162 | 220 | 83 | 141.9 |
| ММ 7938 | – | 240 | 133 | 229 | 80 | 180.5 |
| VMN no number | – | 203 | 136 | 185 | 45 | 150.7 |
| MM 3074 | – | 240 | 155 | ~230 | ~150 | 154.8 |
Conclusion
This study sheds light on the morphology of nasal horns in both modern-day and extinct rhinoceros species. Notably, the nasal horn of living rhinoceroses displays a distinct subdivision into two regions along its height. At the base, the horn widens and closely mirrors the shape of the growth zone, while approximately 5.5 to 8 centimeters above the base, it rapidly narrows until becoming smooth. This narrowing corresponds to the strongly melanized and calcified core part of the horn, a pattern also observed in the woolly rhinoceros.
In the case of the woolly rhinoceros, these findings reveal that the nasal horn’s shape at the base closely resembles that of the nasal rugosity area. However, 5 to 10 centimeters above the base, rapid wear and destruction of the peripheral keratin layer result in a laterally flattened and saber-shaped horn, characteristic of many specimens found in the fossil record (Appendix, Figure 1).
The elongation of the core part of the nasal horn in the woolly rhinoceros may serve a functional purpose, particularly in lateral view where a distinct wear facet is observed on the anterior surface. This wear pattern, likely caused by side-to-side strokes, suggests more physically demanding activities such as breaking snow crusts or ice coatings (Fortelius, 1983).
Moreover, the elongated core part of the woolly rhinoceros nasal horn provides resistance to abrasion, preserving functionality over prolonged use. With a narrower core part, the horn might have worn down to a fracture, rendering it non-functional.
Lastly, this study suggests that the cranial features described herein may aid in reconstructing horn shapes in other fossil rhinoceros species. By examining the shape of the rugosity area and additional outgrowths on nasal bones, researchers can infer the shape of the core part of the horn, contributing to a better understanding of rhinoceros evolution and ecology.
References
Antoine, P.O. (2002). The Elasmotheriina (Mammalia, Rhinocerotidae): Phylogeny and evolution (p. 369). Muséum national d’Histoire naturelle.
Antoine, P.O., & Rookmaaker, K. (2013). The first historical record of a rhinoceros in Togo. Mammalia, 77(1), 127-129. https://doi.org/10. 1515/mammalia-2012-0049
Boeskorov, G. G., Lazarev, P. A., Sher, A. V., Davydov, S. P., Bakulina, N. T., Shchelchkova, M. V., Binladen, J., Willerslev, E., Buigues, B., & Tikhonov, A. N. (2011). Woolly rhino discovery in the lower Kolyma River. Quaternary Science Reviews, 30(17-18), 2262-2272.
Brandt, J. F. (1849). Die Rhinocerotis antiquitatis, seu tichorhini, seu Pallasii structura externa er osteologica observationes, e reliquiis, quae in museis Petropolitanis servantur erutae. Mémoires de l’Académie Impériale des Sciences de Saint‐Péterbourg, Sixième série: Sciences naturalles, 5, 161-416.
Chernova, O. F., & Kirillova, I. V. (2010). New data on horn morphology of the woolly rhinoceros (Coelodonta antiquitatis Blumenbach, 1799). Proceedings of the Zoological Institute RAS, 314(3), 333-342.
Chernova, O. F., Sher, A. V., & Garutt, N. W. (1998). Morphology of the horns of the woolly rhinoceros (Coelodonta antiquitatis). Zoological Journal, 77(1), 66-79.
Eichwald, E. (1835). De pecorum et pachydermorum reliquiis fossilibus, in Lithuania, Volhynia et Podolia repertis. Verhandlungen der Kaiserlichen Leopoldinisch‐Carolinischen Akademie der Naturforscher, 17(2), 677-760.
Ferreira, S., Hofmeyr, M., Pienaar, D., & Cooper, D. (2014). Chemical horn infusions: a poaching deterrent or an unnecessary deception?. Pachyderm, 55, 54-61.
Fortelius, M. (1983). The morphology and paleobiological significance of the horns of Coelodonta antiquitatis (Mammalia: Rhinocerotidae). Journal of Vertebrate Paleontology, 3(2), 125-135.
Garutt, N. W. (1998). Neue Angaben über die Hörner des Fellnashorns Coelodonta antiquitatis. Deinsea, 4, 25-39.
Groves, C. P. (1972). Ceratotherium simum. Mammalian Species, 8, 1-6.
Groves, C. P., & Kurt, F. (1972). Dicerorhinus sumatrensis. Mammalian Species, 21, 1-6.
Groves, C. P., & Leslie, D. M. (2010). Rhinoceros sondaicus (Perissodactyla: Rhinocerotidae). Mammalian Species, 43(887), 190-208.
Hieronimus, T. L., & Witmer, L. (2004). Rhinoceros horn attachment: Anatomy and histology of a dermally influenced bone rugosity. Journal of Morphology, 260(3), 298.
Hieronymus, T. L., Witmer, L. M., & Ridgely, R. C. (2006). Structure of White rhinoceros (Ceratotherium simum) horn investigated by X‐ray computed tomography and histology with implications for growth and external form. Journal of Morphology, 267, 1172-1176.
Laurie, W. A., Lang, E. M., & Groves, C. P. (1983). Rhinoceros unicornis. Mammalian Species, 211, 1-6.
Lazarev, P. A., Boeskorov, G. G., Tomskaya, A. I., Garutt, N. W., Vasiliev, E. M., Kasparov, A. K., & Rodionov, G. N. (1998). Mammals of the anthropogene of Yakutia (p. 167). YaSC SD RAS Press. Ryder, M. L. (1962). Structure of rhinoceros horn. Nature, 193, 1199-1201.
Wilson, D. E., & Mittermeier, R. A. (2011). Handbook of the mammals of the world, Volume 2: Hoofed mammals. Lynx Ediciones.
Appendix
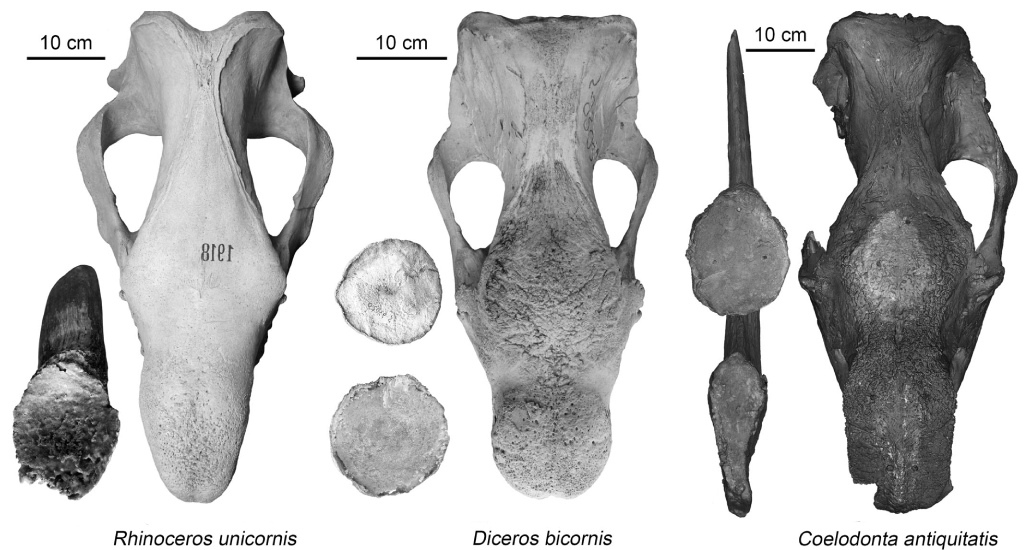
Figure 1: Rugosity area and horn base in woolly rhinoceros species: (a) ZIN 1918; (b) ZMMU S-93020; (c) DPMGI 2114).

Figure 2: Skull with two horns of the Suruktakh woolly rhinoceros (MM 3074).
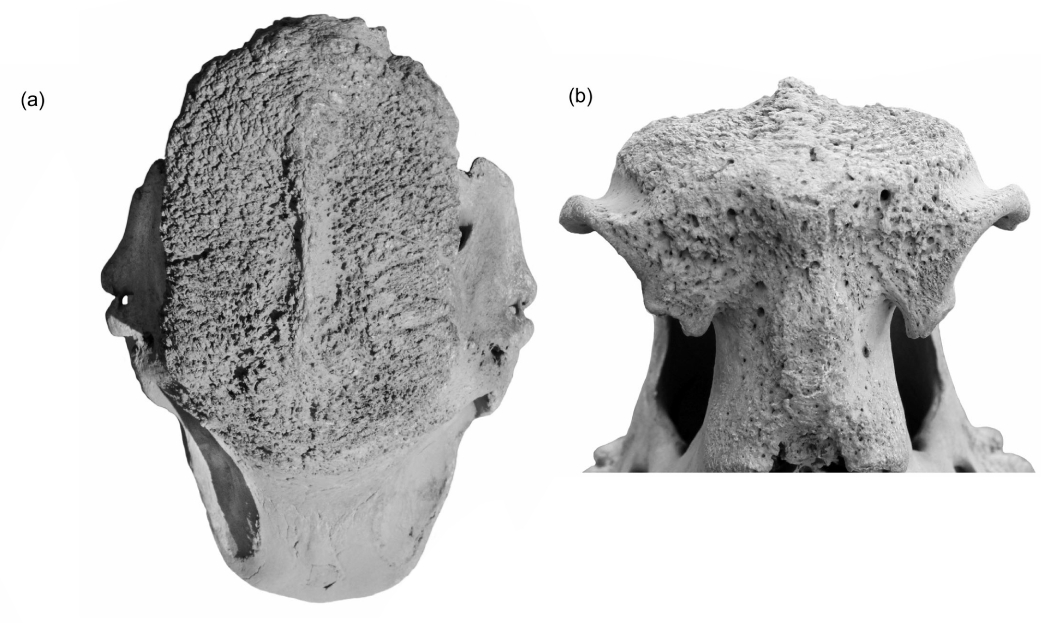
Figure 3: The ridge running lengthwise along the nasal bones of the remains of the Yana River woolly rhinoceros (DPMGI 993).
Sylvia Anderson is a Ph.D. candidate at CAD University, with research focused on the Pleistocene epoch. Concentrating on the geological and ecological aspects of this era, she explores the environmental conditions, biodiversity, and climatic changes that defined the Pleistocene.
